SUNY ESF
Soil Results

By Melany Fisk, Assistant Professor, Miami University of Ohio
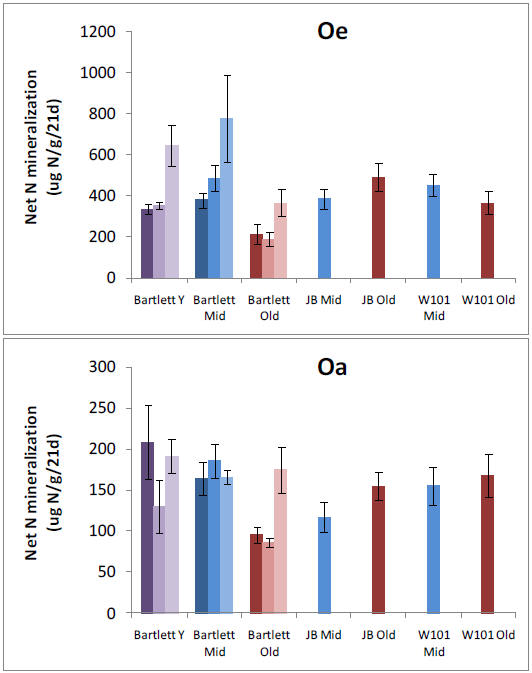
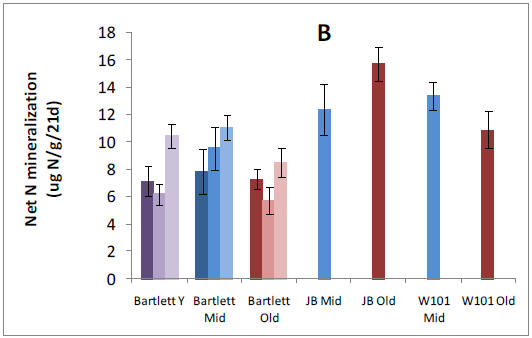
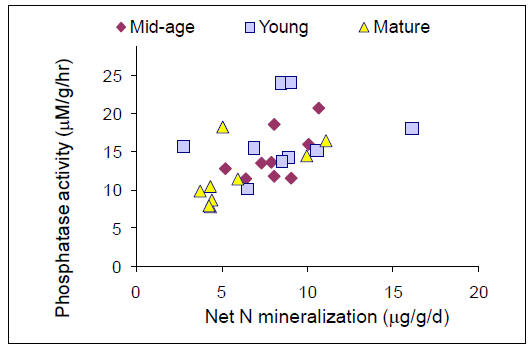
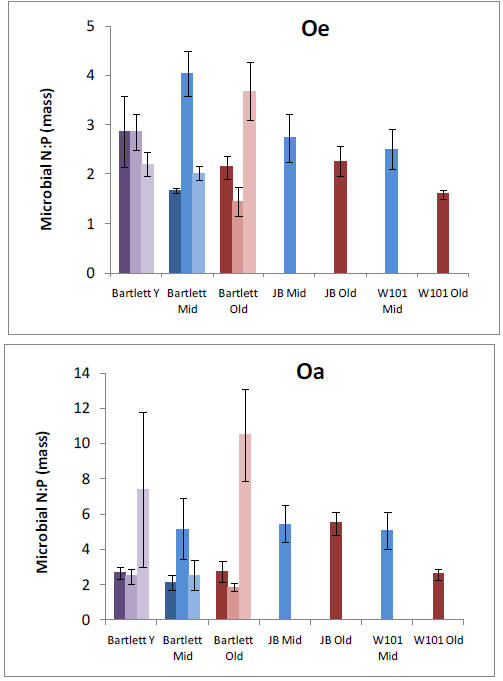
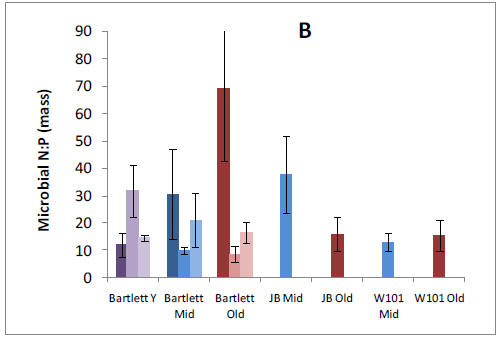
We established plots in 13 forest stands and collected pre-treatment data in preparation for nitrogen (N), phosphorus (P), and NXP fertilization that will begin in May, 2011. We have analyzed patterns of net N mineralization, resin-available P, soil P fractions, soil microbial respiration and microbial N:P ratios across all stands in organic and mineral soil horizons. We tested whether N and P availabilities vary in relation to one another or independently across our 9 replicated forest stands within the Bartlett Experimental Forest. We found a clear positive relationship between N mineralization and resin-P availability that was somewhat surprising given the variation in forest age, species composition, and site characteristics that were encompassed by our study sites. Stoichiometry of the decomposer microbial community might be expected to mediate this process, but we found no relationships between microbial N:P and nutrient availability. However, we did find a strong positive relationship between N mineralization and acid phosphatase activity, suggesting a mechanism by which organisms are using available N to maintain balance with P availability. A relative abundance of organic P in these forests suggests that this resource is potentially available in response to further N enrichment, and we are in the process of analyzing soil P fractions to test whether slowly recycling fractions are depleted in stands with higher N availability.
We have begun pre-treatment analysis of soil fungal communities in our forest stands in order to identify potential functional groups of interest for future studies of hyphal foraging for different nutrient resources. Soils were collected in stands of different ages and varying availabilities of N and P and we are now processing those in the lab.
Findings:
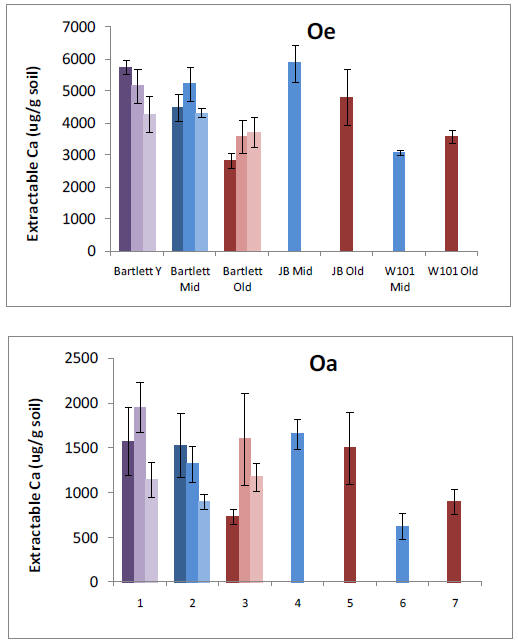
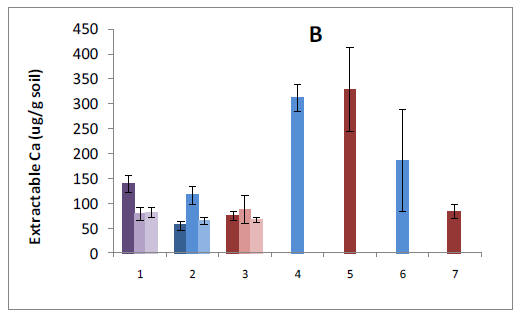
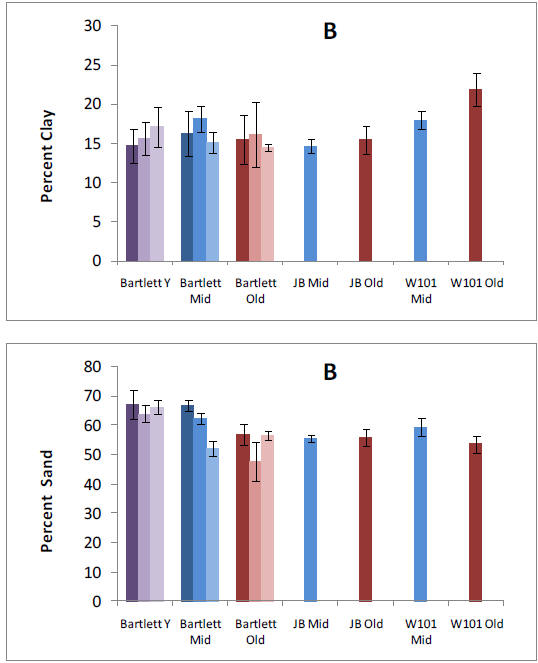
Our pre-treatment work in the Bartlett Experimental Forest has shown that plant roots forage primarily for P or Ca in forest stands where N availability is high. We have also found that allocation to phosphatase tends to increase P availability where N availability is high. These results suggest important mechanisms by which plants and soil organisms allocate effort to balance nutrient acquisition. The lack of correspondence with microbial N:P ratios emphasizes the need to consider turnover through the microbial pool, which may vary in relation to plant allocation belowground. Our results help us to understand spatial variation in biotic processes at the forest-stand level and raise intriguing questions about the sources of that variation. They also provide an important background for testing responses by these processes to changes over time in nutrient availability, as we begin our treatments.